¿POR QUÉ HAY GAYS? La amistad masculina como amor asexuado.
Primero los hechos, empecemos observando la realidad:
LA HOMOSEXUALIDAD: UNA CONSTANTE.
La homosexualidad existe en la especie humana como
una parte minoritaria pero constante de la población humana, en todas las clases,
en todas las culturas, en todas las épocas, la represión de esta habla de su existencia
ubicua.


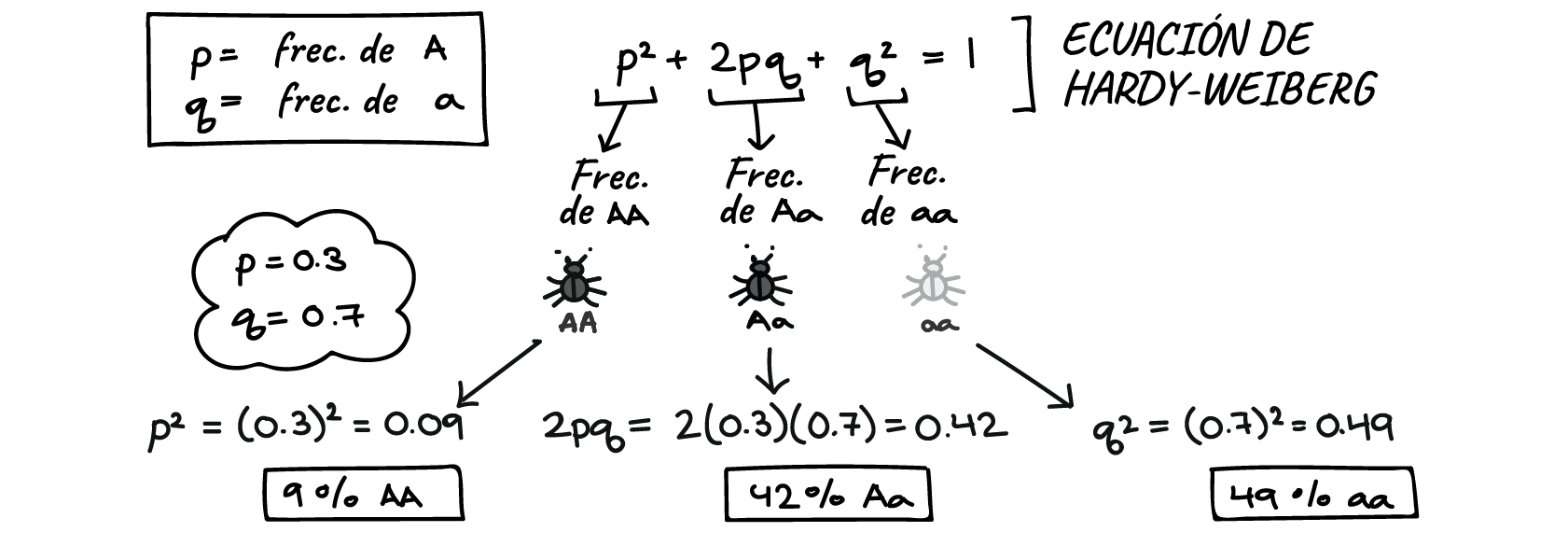
LA PARADOJA DEL GEN GAY
Un gen gay
parece paradójico, pues se presume que este rasgo reduce la fertilidad o el
paso de genes a la siguiente generación. Según la genética de poblaciones (Ley
de Hardy Weiber) solo es posible que un gen permanezca en una población a lo
largo de las generaciones si cumple un rol o tiene un fitness evolutivo
positivo.
Dado que hay
evidencia de herencia genética en la homosexualidad, (Hamer et
al 1993, 1994; Hu et al 1995; Gavrilets et al 2006; Iemmola et
al 2008; Lyons et al 2004; Mustanski et al 2005; Pillard et
al 1982 y 1981; Savolainen et al 2007; Murphy et
al 2005), el llamado
“gen gay” debería servir de algo.
Ya se han
hipotetizado algunos mecanismos de como esto es posible (sin verificación), (Gavrilets
et al 2006; King et
al 2005; MacIntyre et al 1993; McKnight et al 1997; Miller
et al 2000; Muscarella et al 2000; Pattatucci et al 1998; Trivers
et al 1974 y Vasey et al 2007), pero yo
daré una teoría original y nueva.
APOYO MUTUO ENTRE MACHOS
En otras especies los machos se matan entre sí o compiten por las
hembras, difícilmente cooperan o hacen amistad. Pero en el desarrollo de la
especie humana, dado las dificultades ambientales, la fragilidad física del
humano, sobre todo al salir de África, y dada la poligamia femenina, se hizo
menos útil la competencia y se hizo útil la amistad, lealtad y afecto entre
machos humanos, la bioquímica evolucionó para hacer posible el afecto entre
hombres, “friendship has adaptive benefits” (Lauren et al, 2014).

.jpeg)
AMISTAD ≈ AMOR
En los amigos hombres
existe una bioquímica del amor parcial (neurotransmisores semejantes los del
amor, como feniletilamina, serotonina, dopamina, oxitocina etc.).
También las zonas del cerebro
activas en el amor se activase la amistad entre hombres. La neurobioquímica del amor
es casi la misma que la neurobioquímica de la amistad. Quizás la diferencia
solo sea la dosis.

LA AMISTAD, ES COSA DE HOMBRES.
La amistad entre hombres tiene algunos componentes
bioquímicos semejantes al del amor romántico, no así en las mujeres, es fácil
suponer la utilidad de esto evolutivamente hablando, el apoyo mutuo de los cazadores
y guerreros, funciones básicamente masculinas. Esto explica el apego entre
amigos, que incluyen celos, y rituales, como beber justos, al beber baja la
serotonina, justamente este neurotransmisor baja en el amor, así que irse de copas
con los amigos es irse a enamorar un poco de ellos.

LA HIPÓTESIS: UN GEN GAY
PARA EL APOYO MUTUO LLEVO A LA HOMOSEXUALIDAD
Hipótesis:
“La evolución hizo aparecer
la amistad como una variante no sexualizada del amor entre hombres que
necesitaban cooperar, algunos reciben una dosis extra y se sexualiza, dando
origen a la homosexualidad”.
Esto permitió su asociación no sexualizada, o solo
secundariamente sexualizada.
Se puede inferir de esto
que la evolución humana incluyó un poco de “amor” y “erotismo” entre todos los
hombres para vincularlos y acercarlos, de ahí la natural tendencia formar
mejores amigos, pandillas y grupos, este pegamento les permitió salir a cazar
juntos mamuts, o afrontar las guerras con otras tribus.

Los
hombres primitivos usaron este gen gay para unirse y cazar juntos o pelear con
sus cerebros y cuerpos adoptando una forma fisiológica de amor.
LA
EVOLUCIÓN NO CREA EX NIHILO
Ya
sabemos que la evolución no crea de la nada, sino adapta mecanismos previamente
existentes (en este caso la bioquímica del amor) en una función nueva. La evolución
del flagelo es un buen ejemplo. Todas sus partes sirvieron antes para otra cosa.
Nada se creó de novo.
La evolución no crea nada de la nada, generalmente un
nuevo rasgo o función, no es creado desde cero, sino que es una variación o
modificación de un rasgo o función compleja diferente, es decir la evolución
usa como materia prima los órganos estructuras y funciones preexistentes que
tienen otras funciones. Así, las aletas pasaron a ser patas (diferente
función), esto es común en la historia de casi todos los genes.
Podría plantarse así: la
amistad es una variante asexuada del amor.

No hay alguna gente con el gen gay, toda la humanidad
tiene un “gen gay” Este permite la amistad, en algunos casos se ésta se
sexualiza.
EXPLICACIONES, RETRODICIONES
Y PREDICCIONES
Toda teoría debe explicar algo
nuevo, del pasado, del presente y del futuro, debe ser fértil. Aquí resumo algunos nudos que desata esta hipótesis:
.jpeg)
1.-LA
PARADOJA DEL GEN GAY RESUELTA
Todo
gen sirve, el gen o genes gays sirven. Permitieron la amistad que sirvió la
sobrevivencia en entorno adverso, creando una variante asexuada del amor. La
evolución despertó un poco de amor entre hombres, cuando se le pasa la mano
surge lo gay.

2.-NO
UN GEN GAY EN LOS HOMOSEXUALES SINO UN GEN GAY EN TODA LA HUMANIDAD
.jpeg)
Si todos tienen el gen
gay, a veces ese gen es muy exagerado y lleva al sexo. La humanidad está al
borde de la homosexualidad, por eso el tabú a la homosexualidad. El miedo de los
grupos homófobos a que los niños expuestos a imágenes gay se van a “homosexualizar”
es fundado.
3.-LA HOMOSEXUALIDAD ES UN EFECTO LATERAL
Dada la naturaleza poligénica de la bioquímica del amor
(hay muchas zonas del cerebro y moléculas en juego, estas codificadas por multitud
de genes). Es esperable por razones estadísticas una distribución en campana de
Gauss de esta amistad.
.png)
4.- LA HOMOFOBIA SURGE CON LA AGRICULTURA.
Si el gen gay
es común y es útil, ¿por qué la homofobia es generalizada o común en las diferentes
sociedades humanas? ¿Por qué hay homofobia? Esta teoría requiere otra hipótesis,
que acaso solo sea un teorema de la misma premisa:
El modo de obtención de recursos determina la sexualidad
y en resto de la cultura.
Con la agricultura valió más la monogamia heterosexual
y nació la homofobia, pues económicamente, es decir la obtención de recursos, dependía
ahora de otra sexualidad y otra forma de organización social, en las que
poligamia y homosexualidad son reprimidas
La herencia de la tierra por los descendientes genéticos,
es garantizada por la unión monógama de la pareja heterosexual y el control o sujeción
de la sexualidad femenina, antes libre y polígama. Las relaciones homosexuales disminuyen
recursos destinados a la pareja y descendencia que antes la tribu hacía en común.
.jpeg)
5.-LA HOMOFOBIA MUERE CON LAS SOCIEDADES INDUSTRIALES

6.-EL ATAVISMO DE LA HOMOSEXUALIDAD.
El cambio cultural fue brusco,
pero los genes quedaron ahí donde fuertes vínculos entre hombres eran útiles,
persisten como persiste toda esa sexualidad originaria en forma de fantasías,
tabúes, fetiches, imaginarios, juegos y eventuales experiencias y deslices.
7.-HOMOSEXUALIDAD Y NARCISISMO

8.-PREDICCIONES
1.- En la región Xq28 se encontrarán genes asociados a
neurotramisores de amistad o amor.
2.-Las sociedades no agrarias, más cazadoras recolectoras o
descendientes de ellas, hay más gays, como en la amazonía peruana.

Palabras clave
homosexualidad,
genetica, epistemología.
BIBLIOGRAFIA
Gavrilets S, Rice WR. Genetic
models of homosexuality: generating testable predictions. Proc Roy Soc London
B. 2006;273:3031–3038.
Hamer D, Hu S, Magnuson V, Hu N,
Pattatucci A. Male sexual orientation and genetic evidence Science
1993;262:2065.
Hamer DH, Hu S, Magnuson VL, Hu
N, Pattatucci AM A linkage between DNA markers on the X chromosome and male
sexual orientation. Science. 1993 Jul
16;261(5119):321-7.
Hamer DH, Copeland P. The
science of desire: the search for the gay gene and biology of behavior. New
York, NY: Simon & Schuster; 1994.
Hu S, Pattatucci AML, Patterson
C, Li L, Fulkner DW, et al. Linkage between sexual orientation and chromosome
Xq28 in males but not in females. Nature Genetics. 1995;11:248–256.
Iemmola F, Camperio Ciani A. New
evidences of genetic factors influencing sexual orientation in men: female
fecundity increase in the maternal line. Arch Sex Behav. 2008 In press.
Lauren J.N. Brent,1,2 Steve W.C.
Chang,1,2,3 Jean-François Gariépy,1,2 and Michael L. Platt1,2,4. (2014). The
neuroethology of friendship. Ann N Y Acad Sci. May; 1316(1): 1–17.
Lyons MJ, et al. A twin study of sexual
behavior in men. Arch Sex Behav. 2004;33:129–136.
Mustanski BS, Dupree MG,
Nievergelt CM, Bocklandt S, Schork NJ, Hamer DH A genoma wide scan of male
sexual orientation.. Hum Genet. 2005 Mar;116(4):272-8. Epub 2005 Jan 12.
Pillard RC, Poumadere J,
Carretta RA. A family study of sexual orientation. Arch Sex Behav.
1982;11:511–520.
Pillard RC, Poumadere J,
Carretta RA. Is homosexuality familial? A review, some data, and a suggestion.
Arch Sex Behav. 1981;10:465–475.
Savolainen V, Lehmann L.
Genetics and bisexuality. Nature. 2007;445:158–159
Timothy F Murphy The search for
the gay gene BMJ. 2005 April 30; 330(7498): 1033
King M, Green J, Osborn DPJ,
Arkell J, Hetherton J, et al. Family size in white gay and heterosexual men.
Arch Sex Behav. 2005;34:117–122.
MacIntyre F, Estep KW. Sperm
competition and the persistence of genes for male homosexuality. Biosystems.
1993;31:223–233.
McKnight J. Straight science?
Homosexuality, evolution and adaptation. London: Routledge; 1997.
Miller EM. Homosexuality, birth
order, and evolution: Toward an equilibrium reproductive economics of
homosexuality. Arch Sex Behav. 2000;29:1–34
Muscarella F. The evolution of
homoerotic behavior in humans. J Homosex. 2000;40:51–
Pattatucci AML. Molecular
investigations into complex behavior: lessons from sexual orientation studies.
Hum Biol. 1998;70:367–386.
Trivers RL. Parent-offspring
conflict. Am Zoo. 1974;14:249–264.
Vasey PL, VanderLaan DP. Birth
order and male androphilia in Samoan fa'afafine. Proc Roy Soc London B.
2007;274:1437–1442.
Kinsey AC, Pomeroy WR, Martin
CE. Sexual Behavior in the Human Male. Philadelphia: Saunders; 1948
Rice G, Anderson C, Risch N,
Ebers G. Male homosexuality: absence of linkage to microsatellite markers at
Xq28. Science. 1999;284:665–667.
posted by Luis Arbaiza at 11:37 p. m.
0 comments
![]()
![]()

